
音声(発声および聴覚)の研究

発声反応のオペラント条件づけを用いて、動物の音声研究を行うため、小鳥用実時間音声処理systemを開発しました。これまで、動物の発声反応のオペラント条件づけを行うためには、実験者が耳で聞いて、その発声が望ましいものかどうか判断しなければなりませんでした。このsystemの開発によって、特定の発声を行ったとき、あるいは、その発声と異なる発声を行ったときに客観的にそして即座に、強化子(餌など)を提示することが出来るようになりました。このsystemは明星大学・Duke大学・Maryland州立大学・日本大学(生物資源)に設置されました。
アメリカ心理学会News Letter “Monitor”での紹介記事
以下の研究は、そのsystemを用いた研究と、ヒトの聴覚および視覚と聴覚の相互作用に関する研究です。

Manabe, K., Kawashima, T. and Staddon J.E.R. (1995). Differential Vocalizations in Budgerigars: Toward an Experimental Analysis of Naming. Journal of the Experimental Analysis of Behavior, 63, 111-126.
本実験場面の動画は、本HPのビデオライブラリーでご覧になれます。
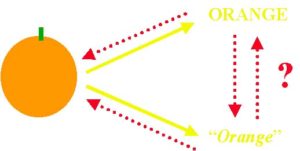
ヒトは事物やモノに対して「命名」することが出来る。例えば、”リンゴ”には音声である/リンゴ/、あるいは、文字としての「林檎」などをあてることが出来る。また、他者が発声した/リンゴ/という音声や、他者が書いた「林檎」という文字は、”リンゴ”を意味することを理解できる。ヒトの命名反応の特徴は、1)異なる事物には、異なった音声などの異なった反応を行う。例えば、”椅子”には、/イス/、”机”には/ツクエ/という異なる発声を行う。2)事物と命名の間の関係は任意である。例えば、同じ”リンゴ”でも、英語では/apple/で日本語では/リンゴ/である。3)目の前にその事物が無くても、その名前を言うことにより、その事物を指示することが出来る。4)全く物理的にあるいは見かけが異なる事物に対しても、同じ名前を与えることが出来る。例えば、腕につける腕時計も/トケイ/であり、砂時計も/トケイ/である。 本論文では、この様な命名反応の特徴はヒトに特有であるのか、セキセイインコを用いて比較心理学的検討を行った。まず、この実験のために開発した動物用実時間音声処理システムを用い、2種の鳴き声(高い鳴き声と低い鳴き声)を鳴きわける訓練を行った。次に、あるセキセイインコには、赤色が提示されたときは高い鳴き声、緑が提示されたときは低い鳴き声を発声させることを学習させた。別のセキセイインコには、この逆の鳴き声を発声するよう訓練した。ここまでで、ヒトの命名反応の特徴である1)と2)がセキセイインコでも可能であることが確かめられた。 その後、二種の形刺激(○と+)のうちの一方を、上述の二種の色光刺激(赤と緑)の一方に対応づけ、別の形刺激は別の色刺激に対応づけるという訓練を行うと、それぞれの形刺激に対しても、「自発的」に対応づけられた色刺激に対する発声反応を行うようになることを確かめた。これは、ヒトの命名反応の特徴である3)と4)がセキセイインコでも可能であることを示唆している。
関連論文
Sidman, M. (1995). Functional classes and equivalence via three-term contingencies. In M. Sidman (Ed.), Equivalence relations and behavior: A research story (pp. 463-474). Boston, MA: Authors Cooperative, Inc.

Saunders, K. J. and Williams, D. C. (1998). Do parakeets exhibit derived stimulus control? Some thoughts on experimental control procedures. Journal of the Experimental Analysis of Behavior, 70, 321-324. Abstract
山崎 由美子(1999). 動物における刺激等価性 動物心理学研究 49(2), 107-137.
室伏 靖子(1999). ”等価な関係”の新展開-反応の機能について- 動物心理学研究 49(2) 217-228.
Sidman, M. (2000). Equivalence relations and the reinforcement contingency. Journal of the Experimental Analysis of Behavior, 74, 127-146. PDF file Abstracts
Urcuioli, P. J. Pierce, J. N. Lionello-DeNolf, K. M. Friedrich, A. Fetterman, J. G. & Green, C. (2002). The development of emergent differential sample behavior in pigeons. Journal of the Experimental Analysis of Behavior, 78, 409-432. PDF file Abstract
Dooling,R.J., Brown, S.D., Manabe,K., and Powell,E.F. (1996). The perceptual foundations of vocal learning in budgerigars. In Moss, C.F. and Shettleworth, S. (Eds.) Neuroethological Studies of Cognitive and Perceptual Processes, pp. 104-128.

ヒトでは、外国語の学習のように、一生涯にわたって音声や発声の学習が可能であるが、多くの鳥では歌や地鳴きの学習がある時期に限定される。しかし、オウムやキュウカンチョウなどは生涯にわたって発声の学習が可能である。本章では、ヒトの音声学習により近い発達過程を持つと思われるオウムの一種であるセキセイインコの音声学習、音声発達、聴覚、視覚を検討した実験研究を概説した。その中で、地鳴き学習はオス・メス共に行い、個体間の社会的相互作用により生涯にわたって修正・学習されることを示した。また、その基礎として聴覚・視覚の特性を実験的に明らかにした。

Manabe, K., Staddon J.E.R. and Cleaveland J.M. (1997). Control of Vocal Repertoire by Reward in Budgerigars. Journal of Comparative Psychology, 111, 50-62.

Nバック手続きを用いて、様々な発声反応が分化強化可能であるかどうかセキセイインコを用いて検討した。このNバック手続きは、直前のN種の反応とは異なる反応を行ったときにのみ強化する(餌を与える)ことにより、反応の変動性を高める手続きである。Nの値を漸次増加していくと、N+1個の発声反応が分化した。スペクトログラムと多次元尺度構成法を用いて発声反応の分化過程を分析したところ、これまで発声していた発声反応の構成要素の組み替え等による「新たな」発声反応の出現が見いだされた。

Manabe, K. and Dooling R.J. (1997). Control of Vocal Production in Budgerigars (Melopsittacus undulatus). Selective Reinforcement, Call Differentiation, and Stimulus Control. Behavioural Processes, 41, 117-132.
発声反応の選択的強化・反応分化・刺激性制御の方法を開発した。これにより、任意の発声反応を任意の時間や刺激により制御することが出来るようになり、その応用として1)人為的難聴などの手法による聴覚と発声との相互作用の研究、2)破壊実験等による発声に関与する脳機能の研究、3)発声反応による命名反応等の認知機能に及ぼす効果の研究等が可能になった。

Manabe, K. (1997). Vocal Plasticity in Budgerigars: Various Modifications of Vocalization by Operant Conditioning. Biomedical Research, 18 ( Supplement 1), 125-132.
発声の訓練方法を、1)反応形成、2)分化強化、3)選択的強化の順に紹介した。2)の分化強化の方法では、徐々に2種類の発声反応に近づけていく方法と、直前に発した発声と異なる発声のみに正のフィードバックを与えることによって複数の発声反応を形成する方法を紹介した。3)では、発声反応の周波数変調の訓練方法と、音量を変化させる訓練方法について概説した。これらの方法は、聴覚障害者の発声訓練にも応用可能である。

Dooling, R. J., Ryals, B. M. and Manabe, K. (1997). Recovery of Hearing and Vocal Behavior after Hair Cell Regeneration. Proceedings of the National Academy of Sciences, USA, 94, 14206-14210.
Abstract Reprint(PDF) Full Text(HTML)
概してヒトやほ乳類の場合、内耳の中の音振動を神経の発火に変換する外有毛細胞が、長期の強い騒音や病気や薬物で一旦死滅すると再生することはなく、生涯難聴に陥る。抗生物質の一種であるカナマイシンも、そのような難聴をヒトやほ乳類では引き起こす。しかし、鳥類の場合、一旦外有毛細胞が死滅した後も再生することが知られている。将来、この再生のメカニズムが解明され、ヒトの難聴の治療に応用されるようになるためには、その再生した外有毛細胞が、正常な死滅前の外有毛細胞に比べてどの程度機能的に回復するものであるか事前に知る必要がある。本論文では、カナマイシンで死滅し、再生した外有毛細胞の聴覚特性および、死滅以前と死滅・回復過程での発声の正確度を測定した。聴覚は、外有毛細胞の死滅に伴って機能が低下し、著しい難聴に陥るが、外有毛細胞の再生に伴って回復することが確かめられた。また、発声の正確度も外有毛細胞の死滅に伴って低下するが、外有毛細胞の再生に伴い、回復することが確認された。ただし、聴力は完全に復活するのではなく、ある程度低下することが確かめられた。

Manabe, K., Sadr E. I. and Dooling R. J. (1998). Control of Vocal Intensity in Budgerigars (Melopsittacus undulatus): Differential Reinforcement of Vocal Intensity and the Lombard Effect. The Journal of the Acoustical Society of America, 103, 1190-1198.
ヒトは、うるさい環境下での会話では、自然(無意識)に発声の音量が大きくなり、静かな環境下では発声の音量は低下する。これは、ランバート効果と呼ばれ、ヒトが環境の音量にあわせて、自分の声の音量を自然にあるいは無意識に調整していることを意味している。本論文では、セキセイインコを被験体として、ヒトと同じようにランバート効果が生じることや、任意に発声の音量を変化させることができることを報告した。
(写真:セキセイインコの鳴き声の音圧を測定するために作成したマイクロフォン付きFM発信器。音圧は音の発生源とマイクロフォンの距離の二乗に反比例して変化する。被験体であるセキセイインコは実験箱の中で動き回るので、マイクロフォンを正面パネルに固定していると、一定の音圧で発声していても、測定された音圧は変化してしまう。従って、この距離を一定にする必要があった。距離を一定にするには、セキセイインコの頭部にマイクロフォンを取り付ける必要があった。しかし、ケーブルがついていると重いため、超小型軽量のマイクロフォン付き発信器を作る必要があった。そこで、コウモリのために開発された超小型FM発信器を参考に、厚さ3mm横8mm縦が10mmで重さは約1gのものを作成した。発信器は、電子回路とマイクロフォン、電池および白金線のアンテナで構成されていた。マイクロフォンおよび電池は、補聴器用のものを転用した。)
真辺 一近・河嶋 孝・J.E.Miller・R.J.Dooling (1999). コンピュータを用いた音声知覚の測定:日本人の/ra/-/la/音の知覚と聴覚障害児(補聴器・人工内耳装用児)の英語の母音の知覚 行動分析学研究, 13,100-114.

日本人は、英語の/s/-/th/や /ba/-/va/等と同様に、/ra/-/la/音の聞き分けが困難であることが知られている。この/ra/と/la/音の物理的な違いは、第3フォルマントにあり、/ra/音では周波数が上昇するのに対して、/la/音では変化しない。この第3フォルマントの周波数軸と時間軸上での傾きを徐々に変えることにより、/ra/から/la/音に徐々に変化する一つの音の連続体を作ることが可能である。英語がネイティブである被験者にそれらの合成音を/ra/あるいは/la/に分類させた場合、物理的には徐々に変化しているにもかかわらず、傾きがある一定以上であればほとんどの場合/ra/音に分類され、それ以下であればほとんど/la/音に分類されるというカテゴリカルな知覚をしていることが知られている。これに対して一般の日本人では、このようなカテゴリカルな知覚は存在しないことが知られている。 本論文では、コンピュータを使用して、日本人の/ra/-/la/音の知覚の様相を測定する方法を発表した。本来ネイティブが手がかりとしている、第3フォルマント以外の変数(例えば音圧)を変化させるなどの操作を加えると、ネイティブはほとんど影響を受けないのに対して、日本人の反応は大きく影響を受けることを確かめた。従って、特にネイティブでない被験者(被検者)や、健聴者と異なる聴覚を持つ被験者の音声知覚の測定を行う場合、目的とする変数以外の変数を相殺するような手続きを導入する必要があることが確かめられた。また、音の相違の検出に要する時間(反応潜時)を利用して、ネイティブとの知覚の類似度を多次元尺度構成法により算出し、その被験者の「ネイティブ度」を尺度化する事が可能であることを示した。さらに、この手法は、人工内耳装用児や、補聴器装用児などの聴覚に障害を持つ児童の聴能測定にも有効であることが示された。
関連論文
長田 佳久 (1999). 真辺 一近・河嶋 孝・J.E.Miller・R.J.Dooling論文を読んで 行動分析学研究14, 15-16.
批評論文へのreply
真辺 一近・河嶋 孝 (1999). 長田氏のコメントへのリプライ 行動分析学研究 14, 17-19.
Manabe, K. and Riquimarox, H. (2000). Sound controls velocity perception of visual apparent motion. The Journal of Acoustical Society of Japan (E), 21, 173-176.
聴覚刺激と同時に視覚刺激を提示すると、聴覚刺激の知覚が変化する場合があることが報告されている。例えばマガーク効果では、/ba/という音声と同時に、/ga/という音声を発声するときの唇の動きをビデオで提示すると/da/と言ったように聞こえる。この論文ではこれとは逆に、聴覚刺激の提示が視覚刺激の知覚に影響を与える現象を見いだし、それを報告した。光点を順次点滅していくと、光が移動しているように見える「仮現運動」が知覚される。このとき、短い音を提示すると、光点の動きの速さが、音を提示しない場合、あるいは相対的に長い音を提示した場合に比べて、より速く知覚されるというものであった。音の提示時間の効果以外に、音圧の効果、音を提示する耳側(ラテラリティ)の効果、音の提示のタイミング等についても検討した。その結果、音の提示時間に比べて、音圧・ラテラリティ・音の提示のタイミングなどの効果は比較的小さいことが確かめられた。次に、本実験場面において、聴覚と視覚の時間分解能を測定したところ、聴覚の方が優れていることが確認された。現在の時点では、どの様なメカニズムで提示された聴覚刺激が視知覚における速さ知覚に影響を与えるのか、まだ解明されていない。この問題は今後の課題である。
Manabe, K., Dooling, R. J., and Brittan-Powell, E. F. (2008). Vocal learning in Budgerigars (Melopsittacus undulatus): Effects of an acoustic reference on vocal matching. The Journal of the Acoustical Society of America, 123(3), 1729-1736.

Abstract Reprint(PDF) Full Text(HTML)
セキセインコの発声反応をオペラント条件づけにより強化した。まず、コンピュータにデジタル記録された鳴き声(ターゲットコール)と同様な鳴き声を発声したときにのみ強化することにより、特定の鳴き声を分化強化した。十分正確に発声するようになった後、その鳴き声を試行の直前に聞かせた場合の発声の正確度と発声の強度に与える効果について吟味した。その結果、正確度および強度が上昇した。一方、ターゲットコールの提示から、発声するまでの潜時を分化強化によって制御した場合には、正確度および発声の強度は影響を受けなかった。次ぎに、ターゲットコールを、別の鳴き声に変化させ、漸次的接近法により訓練を行うことにより、発声反応が変化することが示された。実験箱の中で訓練された鳴き声は、主として実験箱の中で鳴き、実験以外の時間すごしている飼育室の中ではほとんど鳴くことはなかった。
シンポジウム・講演など
1. 平 4. 7 ラウンドテーブル「行動分析における音声処理システムの利用」話題提供者 日本動物心理学会第52回大会
2. 平 9. 9 小講演「発声反応のオペラント条件付けとその応用:動物から人へ」 日本心理学会第61回大会
3. 平10.7 ニューロクラブセミナー「小鳥の鳴き声の条件付けとその応用:認知研究から神経生理まで」話題提供者 名古屋大学先端科学研究センター